生態圖文culture
殼狀珊瑚藻專區
基本介紹
殼狀珊瑚藻在分類上歸屬於海洋性紅藻珊瑚藻目,為一群古老的海洋紅藻,廣泛分佈於世界各海洋的珊瑚礁海域和岩岸地形。因具行光合作用能力,在生長時亦需要適當的水溫、光照及營養鹽。
過去研究紀錄顯示臺灣北部沿海以殼狀珊瑚藻為主體的藻礁分佈,從新北市石門洞、富貴角、麟山鼻、三芝與淡水、桃園市觀音至新竹縣新豐,皆有大面積的藻礁,其中以桃園觀音藻礁(=礁灰岩)面積為最大。

1維石藻未確定種

2張伯倫氏藻未確定種

3G1 區藻礁現況_1
藻礁的形成:
殼狀珊瑚藻或稱造礁珊瑚紅藻會分泌鈣質或矽質骨骼,經過漫長時間的堆積而逐漸形成藻礁。
藻礁的形成:
殼狀珊瑚藻或稱造礁珊瑚紅藻會分泌鈣質或矽質骨骼,經過漫長時間的堆積而逐漸形成藻礁。

4G1 區藻礁現況_2

5G2 區藻礁現況_1

6G2 區藻礁現況_2

7波緣澎石藻

8澎石藻未確定種

9調查照
潮間帶的小小居民:鰕虎
在大潭藻礁區的潮池中,有時可以發現一群體型嬌小的魚類:鰕虎。鰕虎是魚類中成員最多的科之一,目前已知超過2,000種。共同特徵是腹鰭演化成吸盤,常會攀附在礁岩上,刮食藻類或各類水中有機碎屑。大潭藻礁區可以發現多種鰕虎,像是黑深鰕虎、康培氏銜鰕虎、藍點深鰕虎等。牠們的花色五彩斑斕且多樣性高,可以很好的與環境融為一體。如有機會退潮時來到藻礁區,不妨找找這些潮間帶的小小居民。

1圖1:康培氏銜鰕虎

2圖2:黑深鰕虎

3圖3:藍點深鰕虎
殼狀珊瑚藻點點名
形成大潭藻礁區的殼狀珊瑚藻種類繁多,它們皆屬於紅藻門珊瑚藻亞綱下的物種。在行光合作用時,殼狀珊瑚藻會將海水中的碳酸氫根離子轉換成碳酸鈣沉積,經年累月形成藻礁礁體。形成礁體的礦化作用不但能控制海水酸化,也有利於形成多樣化的生物棲地。在大潭藻礁區進行生態調查時,判斷珊瑚藻種類是重要但難度不低的工作。下面有幾張不同種類珊瑚藻的相片,讀者可以試看看能否分辨其中差異。

1圖1:孢石藻

2圖2:玫瑰哈維石藻

3圖3:太平洋殼葉藻
美麗與美味兼具:玳瑁石斑
在藻礁區的魚類資源調查中,時常可以發現一種帶有網紋的美麗魚類。牠是玳瑁石斑,因為花紋長得很像玳瑁(海龜)的背甲而得名。玳瑁石斑廣泛分布於臺灣沿海潟湖或礁岩區,體長普遍不到30公分,以小魚、甲殼類為食。同為石斑,玳瑁石斑常為大潭在地居民所捕捉利用;因為其美味且與民眾生活緊密相依,被澎湖縣選為縣魚。台灣中油致力於藻礁保育,透過維持良好的棲地環境,讓玳瑁石斑等經濟魚種繁衍生息,對漁業資源永續利用產生正面助益。

1圖1:魚類資源調查時捕獲的玳瑁石斑
橫行霸道的盔甲武士:兇猛酋婦蟹
在藻礁區中,常可以見到一種全身呈深紅色,連眼睛都是紅色的螃蟹在礁岩中爬行。這是「兇猛酋婦蟹」,是在岩質潮間帶中常見的掠食者,以貝類等物種為食。顧名思義,其性格相當兇猛,搭配強而有力的大螯,讓人望而生畏。在進行生態導覽時,民眾最好奇的就是此種螃蟹究竟能否食用?要提醒的是,兇猛酋婦蟹會累積一些自進食而來的毒素,所以並不建議食用。有機會來到藻礁區,不妨花點心思找找兇猛酋婦蟹的身影吧。

1圖1:緊盯著鏡頭的兇猛酋婦蟹

2圖2:躲藏在藻礁複雜結構中的兇猛酋婦蟹
在藻礁活動的水鳥朋友
擁有沙地、泥灘及礁岩等豐富地貌的藻礁區,對水鳥們而言可說是高檔自助餐廳。每年冬季,總會吸引許多冬候鳥拜訪大潭藻礁,並在此度冬及繁殖。其中數量最多的是東方環頸鴴、太平洋金斑鴴等鴴科鳥類,這些體型嬌小的水鳥常會集體棲息在沙地或藻礁上,退潮時集體飛至潮間帶覓食。鴴科鳥類主要以昆蟲、軟體動物、甲殼類動物等小型無脊椎動物為食,覓食時會先跑動一段距離,停下啄食後再繼續跑至下一點,動作十分敏捷逗趣。有機會來到藻礁區,不妨觀察這些水鳥朋友的身影;但為了避免干擾,記得保持距離眺望或借助望遠鏡欣賞喔。

1圖1:成群結隊的太平洋金斑鴴

2圖2:在沙地上停棲的東方環頸鴴

3圖3:在泥灘地上覓食的東方環頸鴴
嬌小的捉迷藏高手:潮間帶中的小型蟹類
在藻礁的岩縫裡,時常可以見到小型蟹類在其中活動。這些蟹類中有相當數量是方蟹總科的成員,包括小厚紋蟹、小型小相手蟹、平背蜞等,體型介於數公釐到5公分之間。雖然體型嬌小,但牠們擁有多變的體色,從黑褐色、螢光綠、紫紅色到紅棕色都有。豐富的色彩並不只是好看,還讓這些蟹類可以與粉紅色的殼狀珊瑚藻、綠色的大型藻類和灰褐色的藻礁礁體融合在一起;配合敏捷的移動速度,可說是藻礁區的捉迷藏高手。要找到並辨識擁有保護色且動作迅速的迷你蟹類,是很需要經驗及技巧的。
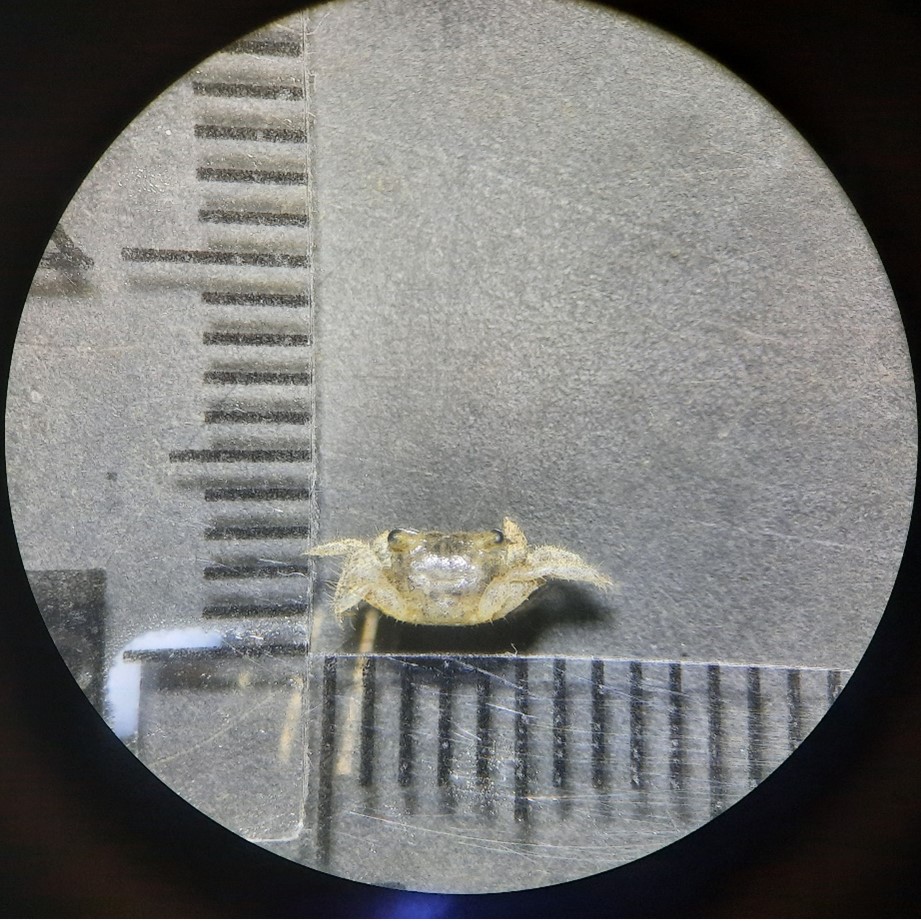
1圖1:小厚紋蟹

2圖2:小型小相手蟹

3圖3:平背蜞
熟悉又陌生的居民:在藻礁生活的螺類們
走入藻礁區,最容易見到的生物,應該是附著在礁岩或藏在潮池裡的螺類們了。雖然很常能見到螺類,不過除非有特別研究,否則要辨識這些小生物還頗有難度。成群結隊、相依偎一起的黑瘤海蜷,是最為常見的藻礁動物;會捕食牡蠣的蚵岩螺,讓蚵農頭痛不已;附著在岩礁的美珠翼法螺,是藻礁區目前唯一發現的法螺科物種。在過往螺類是在地居民常會食用的對象,隨著時代變遷採集的人漸稀,這些藻礁居民們仍靜靜的在潮間帶繁衍生息。有機會不妨彎下腰,找找這些躲藏在潮池和岩礁中的小生物吧。

1圖1:黑瘤海蜷

2圖2:蚵岩螺

3圖3:喜歡成群結隊的黑瘤海蜷
低調又美麗的古老生物:海綿
藻礁區的生物項,隨著季節與環境變化而呈現不同的樣貌。2024年9月的藻礁生態調查中,除了殼狀珊瑚藻、兇猛酋婦蟹等物種外,我們還發現相當數量的海綿。海綿是海綿動物門物種的總稱,是構造相當簡單且原始的動物,也是海龜、海蛞蝓等海洋生物的食物。海綿沒有消化和循環系統,依靠流過自身的海水獲取食物、氧氣並帶走代謝產物,連帶過濾水中的顆粒和污染物,有助於維繫良好的海洋環境。海綿的分佈範圍相當廣,有機會到礁岩為主的潮間帶,記得試試看找尋牠們的蹤跡。

1圖1:在大潭藻礁發現的蜂海綿

2圖2:於潮池中發現的兇猛酋婦蟹
夏末秋初,繁忙的藻礁生態系
時序已經來到8月底,G3區的小燕鷗也已經啟程南返,即便如此,藻礁區還是可以零星觀察到南返的小燕鷗陸續經過,藻礁區的生物們也依然活耀著。上潮位的沙灘上,角眼沙蟹正在挖洞;沙灘上刺球狀的濱刺麥穎果果實也成熟了,隨著海風四處滾動。艷陽下的上潮位藻礁佈滿了整片綠色的滸苔,黑瘤海蜷跟他的同伴們也聚集在退潮的藻礁區中。時至傍晚,太平洋金斑鴴、東方環頸鴴也飛來藻礁區休憩,潮池裡躲藏的短槳蟹正伺機準備捕捉牠的美味晚餐,藻礁生物們正忙著各自的生存大計。

1圖1:潮池裡的角眼沙蟹往上翹的眼睛角芽露出水面相當逗趣

2圖2:沙灘保齡球-濱刺麥的長刺球狀穎果已經成熟

3圖3:東方環頸鴴來到藻礁區停棲逗留
藻礁中的嬌客:桃園紅足軟珊瑚
在藻礁區,除了殼狀珊瑚藻和大型藻類外,還有許多生物棲息在其中,桃園紅足軟珊瑚是其中之一。桃園紅足軟珊瑚(Erythropodium taoyuane)是紅足軟珊瑚屬家族的成員,為臺灣特有種,模式標本就是在觀塘海域採集的,目前也僅在觀塘藻礁區低潮線處可以發現。在藻礁區進行生態調查時,有時可以在潮池中發現牠們的嬌小身影。以膜狀組織相連的珊瑚蟲依附在藻礁上,隨著潮水晃動的身姿相當美麗。若有機會前來藻礁區,不妨來尋訪此種漂亮的軟珊瑚,要找到牠們還頗需運氣呢。

1圖1:桃園紅足軟珊瑚

2圖2:殼狀珊瑚藻、大型藻和桃園紅足軟珊瑚
中油與保生社區的夥伴關係-藻礁漂沙監測
海岸的沙隨著季節,冬天的東北季風、夏季的西南風,又有時颱風侵襲,都會將藻礁的沙帶來帶去。在三接工程進展之下,為了化解外界對外推方案的三接是否會影響漂沙覆蓋藻礁,中油公司特地央託最懂藻礁的一群在地人-保生社區,共同來監測漂沙的一舉一動。
社區居民習於在崎嶇的藻礁移動,並出動沙灘車協助運送測沙桿,有在地社區協助,很快就將測沙桿設置完成,將可更精細瞭解漂沙的季節變動與對藻礁的影響。所幸有當地社區居民願意挺身而出,與中油站在一起,合力守護這片世居的家園。




紅綠大對抗,藻礁區的海藻競爭
紅色一方是殼狀珊瑚藻,有著粉紅色澤,藉由死亡後的礦物質細胞壁堆疊出藻礁岩體,是形成藻礁最主要的藻類;而代表綠色的一方,則是具有鮮翠綠色的大型藻。以石蓴為主的大型藻,每年4月從北部知名的老梅綠石槽分布到桃園的藻礁區,著生在藻礁上綠意盎然,充滿春天的氣息,但在背後卻上演著一齣不為人知的生物競爭。因為東北季風漸緩海水漸清、海水溫度緩緩上升,環境慢慢變得更加適合大型藻生長,使得原先2、3月因殼狀珊瑚藻而一遍粉紅的藻礁區逐漸轉綠,這種生物競爭年年上演,也構成了藻礁區豐富多樣的生態風貌。




藻礁,生生不息!
歲末年初,大潭海岸幾波寒流,氣溫水溫已降,形成藻礁的造礁藻類-殼狀珊瑚藻逐漸可以明顯觀察到,漸漸將藻礁點綴出淺淺的一抹粉紅,正式宣告殼狀珊瑚藻將開始大量生長。 或許會有疑問,那夏天不會有殼狀珊瑚藻嗎?其實,海洋中有千百種藻類,每種藻類對環境需求不同,在氣候合宜時附著於礁岩生長,不適合時以配子體的形式漂浮在大海等待生長季節的到來。大潭藻礁夏天時,以石花菜及刺腔藻等大型藻為優勢。現在,喜歡冬季的殼狀珊瑚藻接力來營造藻礁生態。


早安。藻安
時序很快的準備進入冬季了,氣溫也逐漸下降,這時候恰好是形成藻礁的殼狀珊瑚藻開始旺盛生長的時節,今天(11/17)趁著大退潮,來到大潭G1藻礁區來看看今年殼狀珊瑚藻著生情形,從上潮帶往中、下潮帶一路前進,可以看到明顯差異,上潮帶一如以往,殼狀珊瑚藻著生較少,而中下潮帶的殼狀珊瑚藻相對較多,已經逐漸附著生長,尤其是在潮池、潮溝處,不管是垂直面還是水平面,都可以看到殼狀珊瑚藻逐漸附著生長。另外,在潮池也看到藻礁區常見的生物,例如兇猛酋腹蟹、短槳蟹、柴山多杯孔珊瑚、寄居蟹等。

1圖一、三接與藻礁

2圖二、造礁藻類:殼狀珊瑚藻

3圖三、兇猛酋婦蟹

4圖四、柴山多杯孔珊瑚
海蛞蝓悠閒漫步於大潭G2藻礁
今(112)年7月13日,調查團隊來到G2藻礁,原本打算使用浮潛方式拍攝柴山多杯孔珊瑚水下的照片,卻意外發現顏色非常鮮豔的幾何角鰓海蛞蝓,看著幾何角鰓海蛞蝓黃色的嗅角及鰓,隨著海流搖曳的舞姿,真的是非常舒壓與療癒呢,大家有空也不妨多多親近大自然吧!

1圖一、水下的幾何角鰓海蛞蝓
海兔悠閒漫步於大潭藻礁
今(112)年5月17日,調查團隊來到大潭藻礁G2區進行柴山多杯孔珊瑚第2季的監測調查工作,在調查過程中意外發現,藻礁區中有許多的睦月瘤砂葵生長,另外還有海兔漫步在現生的殼狀珊瑚藻上面,這個季節剛好是海兔的繁殖季節,下次如果在藻礁看到這些小傢伙,記得不要過度干擾牠們喔!

1圖一、藻礁區的睦月瘤砂葵生長

2圖二、優遊漫步於藻礁上的海兔
112年3月日本絨螯蟹:
112年3月藻礁生態調查時,竟然發現俗稱毛蟹的日本絨螯蟹蹤跡,根據當地居民的描述,毛蟹已經在桃園沿海地區消失許久,看來藻礁的生態在大家的保護下越來越豐富囉!

1圖一、112年3月發現日本絨螯蟹
藻礁小學堂:大潭海灘上這些細碎的顆粒是什麼呢?
(X)是藻礁的屍體
(O)是貝殼破碎的產物
事實上這些白色的細顆粒,是近岸地帶的貝殼與珊瑚礁,在海浪的作用下破碎後,再被海流與潮汐捲帶上岸。由於貝殼質地較輕,往往會覆蓋在其他沙礫的上層,形成一層薄薄白色貝殼沙。這景觀在台灣南北的海灘皆可發現,下次如果到海邊發現白色貝殼沙,可以細看裡面有沒有漂亮的貝殼囉!

1圖一、白玉海岸貝殼沙近照

2圖二、大潭海岸貝殼沙海灘照
111年3月殼狀珊瑚藻:
111年3月,隨著海水溫度下降,殼狀珊瑚藻也恢復活力,慢慢在硬底質上鍍上一層粉紅色的薄膜,吸引貝類、寄居蟹來觀光,兇猛酋婦蟹也跟著來湊熱鬧,整個大潭藻礁區熱鬧了起來。

1圖一、殼狀珊瑚藻與寄居蟹

2圖二、3月殼狀珊瑚藻現況

3圖三、兇猛酋婦蟹

4圖四、沐浴晨光的大潭藻礁
110年5月殼狀珊瑚藻現況:
殼狀珊瑚藻的生長狀況大約到了4月是高峰,之後因天氣炎熱、海水溫度升高,殼狀珊瑚藻的生長將會受到影響,夏季將會白化甚至死亡,死後殼狀珊瑚藻的碳酸鈣骨骼將遺留在礁體上,成為來年新一世代的殼狀珊瑚藻附著生長的基質,年復一年層層堆疊,形成我們現在看到的藻礁。

1圖一、殼狀珊瑚藻近照

2圖二、110/05/27 G1區藻礁現況

3圖三、110/05/27 G2區藻礁現況
藻礁的形成:
殼狀珊瑚藻或稱造礁珊瑚紅藻會分泌鈣質或矽質骨骼,經過漫長時間的堆積而逐漸形成藻礁。

1

2

3

4
殼狀珊瑚藻的世界分布(上):
殼狀珊瑚藻的世界分布全世界由無節珊瑚藻組成的相關地質主要有三種形態,分別是藻礁(Algal reef)、藻脊(Algal ridge = 藻瀑,Algal cascades)與紅藻石床(Rhodolith beds)。
紅藻石床廣泛分布於全球各海域,主要是由非穩定固著於海底基質上的紅藻石所形成的群落。紅藻石床的世界分布紀錄包含東太平洋中部的加利福尼亞灣索諾拉州、南下加利福尼亞州到哈利斯科州之間的小島沿岸、航道、海灣與潟湖等常有波浪及洋流的地區、東北太平洋的英屬哥倫比亞省夏洛特皇后群島、北太平洋阿拉斯加(Herring Bay)、日本琉球、澳洲昆士蘭、法屬玻里尼西亞的茉莉亞島、北大西洋的愛爾蘭沿岸、地中海的西班牙、摩洛哥、阿爾及利亞、法國、義大利、突尼西亞、愛琴海、黎凡特與馬爾馬拉海等地。臺灣也曾在臺東縣東河鄉外海大陸棚上水深約62公尺處,記錄有現生的紅藻球,由表層的無節珊瑚藻與有孔蟲形成包覆形的瘤狀結構。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
殼狀珊瑚藻的世界分布(中):
藻脊又稱為藻瀑,定義為礁體表層具無節珊瑚藻分布,礁體內部則或有其他鈣化生物與岩石共同組成。全世界的藻脊分布於加勒比海的普拉納礁、大伊納瓜島、聖克魯斯島、安地卡島、巴布達島、聖佑達修斯島、馬丁尼克、瓜地洛普、瑪麗-加朗特島、格瑞那丁群島、洛斯羅克斯群島、科蘇梅爾島等地、澳洲西北側塔隆島、巴西的羅卡斯環礁、南太平洋的湯加塔布島等地。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
殼狀珊瑚藻的世界分布(下):
藻礁的礁體大部分由無節珊瑚藻組成,目前於地中海的西班牙托薩德馬爾、義大利、巴利阿里群島、法國的科西嘉、突尼西亞等地、北大西洋的百慕達海底平台記錄有8-12公尺高杯形藻礁及挪威有紀錄。挪威的藻礁記錄於挪威北部特羅姆斯區的外峽灣槽,水深約6-15公尺處海底平台,此處礁體多是由無節珊瑚藻著生於硬基質生物骨骼上後,快速向上生長出樹枝狀個體,接著藻體向上與向水平面增長,再結合不同生長特性的殼狀珊瑚藻後逐漸形成礁體的框架。然而,與臺灣桃園藻礁不同,副北極地區的藻礁因缺乏早期膠結作用的發生且因藻體生長緩慢,在礁體的基部易發生內石性的生物侵蝕,常使礁體的保存受到限制。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
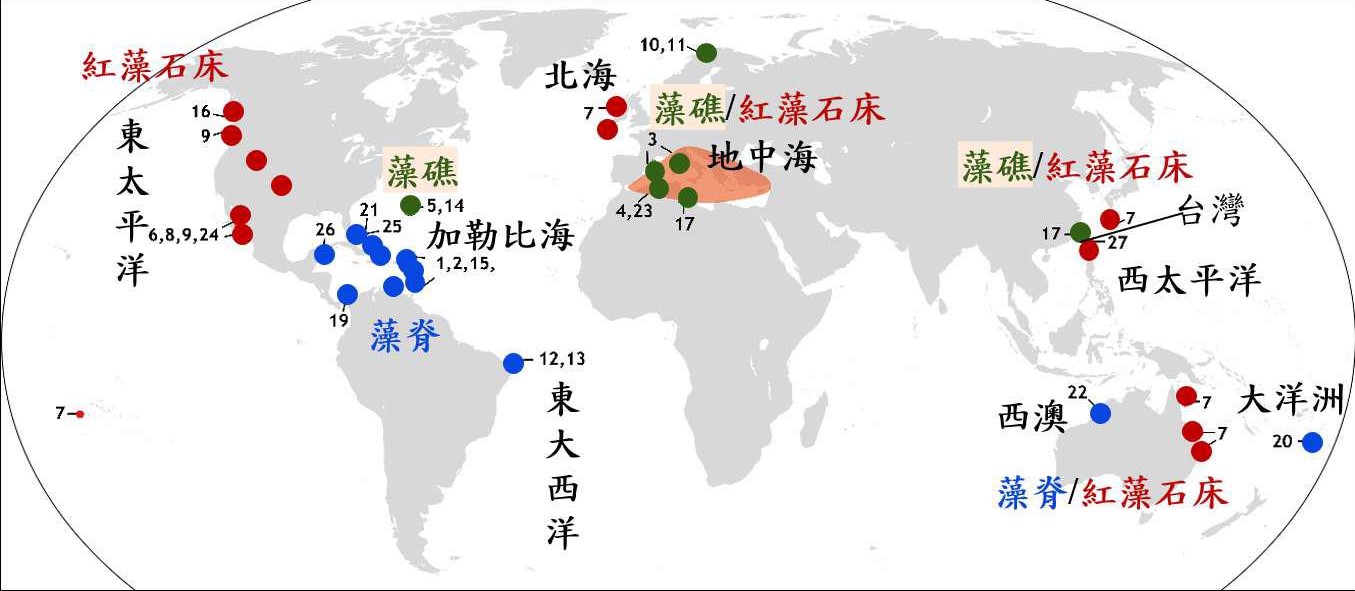
1藻礁(綠色)、藻脊(藍色)與紅藻石床(紅色)的全球紀錄分布圖
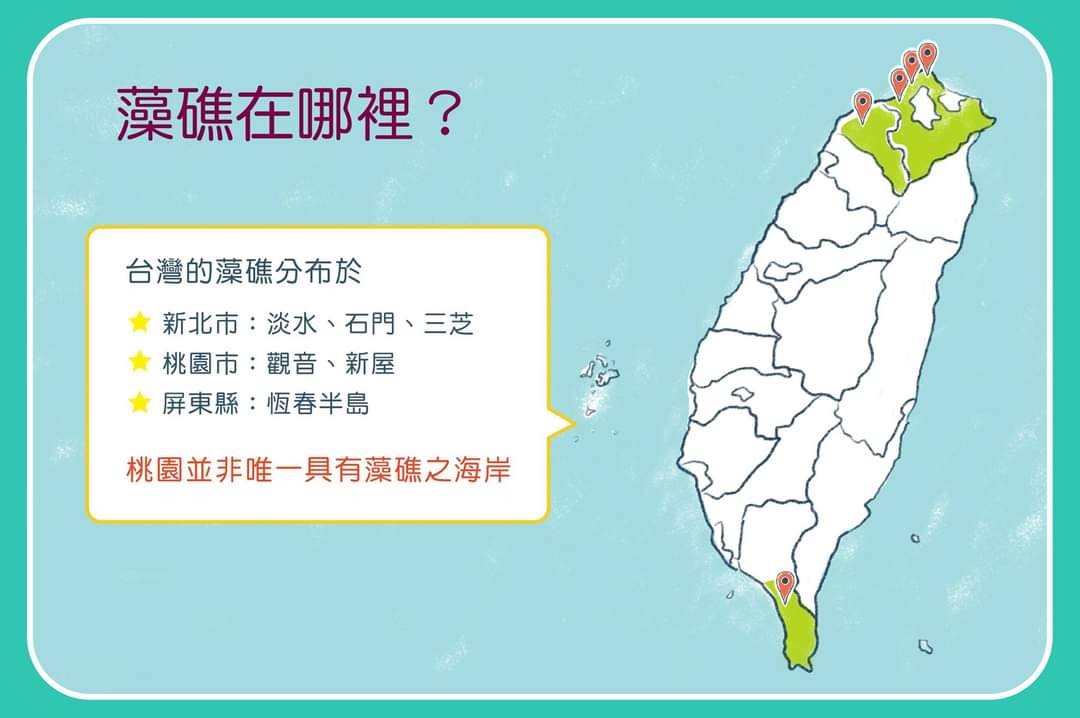
台灣殼狀珊瑚藻的分布:
台灣沿海的生物礁主要為珊瑚礁,分布在南、東、北部和各離島沿岸海域;相對而言,由有活力的殼狀珊瑚藻為主要組成份子的現生藻礁的分布就比較局限。目前臺灣各海域的藻礁分布,除了桃園礁灰岩(=藻礁)海岸外,另有臺北縣石門、三芝鄉與淡水鎮海岸,以及恆春半島的局部海岸(但目前已轉變成珊瑚礁);其中,尤以桃園縣海岸的藻礁分布範圍最為廣泛。
資料來源:
「桃園觀音藻礁生態解說手冊」-戴昌鳳、王士偉、張睿昇、鄭安怡,2009
「礁、藻礁、找藻礁」-王士偉,2010
1
殼狀珊瑚藻生活史(上):
殼狀珊瑚藻的藻種特性為具有鈣化藻體與紅藻特有的三相生活史以及所有生殖結構皆有特化的囊狀結構(conceptacles)保護。三相生活史可分為行無性生殖的四分孢子體世代(二倍體)、有性生殖的配子體世代(單倍體)以及寄生在配子體世代上的果孢子體世代(二倍體)。一般而言,不論是透過無性生殖產生的四分孢子或有性生殖產生的雌雄配子皆在藻體表面的生殖囊腔中發育。在有性生殖的配子體世代是雌雄異體,雄配子體囊腔中的雄配子在發育成熟後由生殖腔上方的囊孔釋放出來,游離的雄配子進入成熟雌配子體的生殖囊腔中與雌配子結合,並在雌配子體生殖囊腔內發育成寄生性的果孢子體世代(二倍體)。
資料來源:
「 110年-111年藻礁生態系調查計畫(2/2)成果報告書」海保署,2022
殼狀珊瑚藻生活史(下):
果孢子體在成熟後,透過雌配子體生殖囊腔上方的囊孔的釋放出果孢子,而果孢子如成功在已形成的藻礁上萌發,則可生長發育為四分孢子體世代。成熟的四分孢子體表面的生殖囊腔中有多個帶狀分裂的四方孢子囊,成熟的四分孢子囊由生殖囊腔上方的囊孔釋放出四個四分孢子,離開囊腔的四分孢子若可順利著生在舊的藻礁上,則可生長發育為下個新世代的雌性或雄性配子體,完成三相世代交替生活史,即為四分孢子體、配子體以及寄生在雌配子體上的果孢子體交替出現的世代交替生活史。
資料來源:
「 110年-111年藻礁生態系調查計畫(2/2)成果報告書」海保署,2022

1
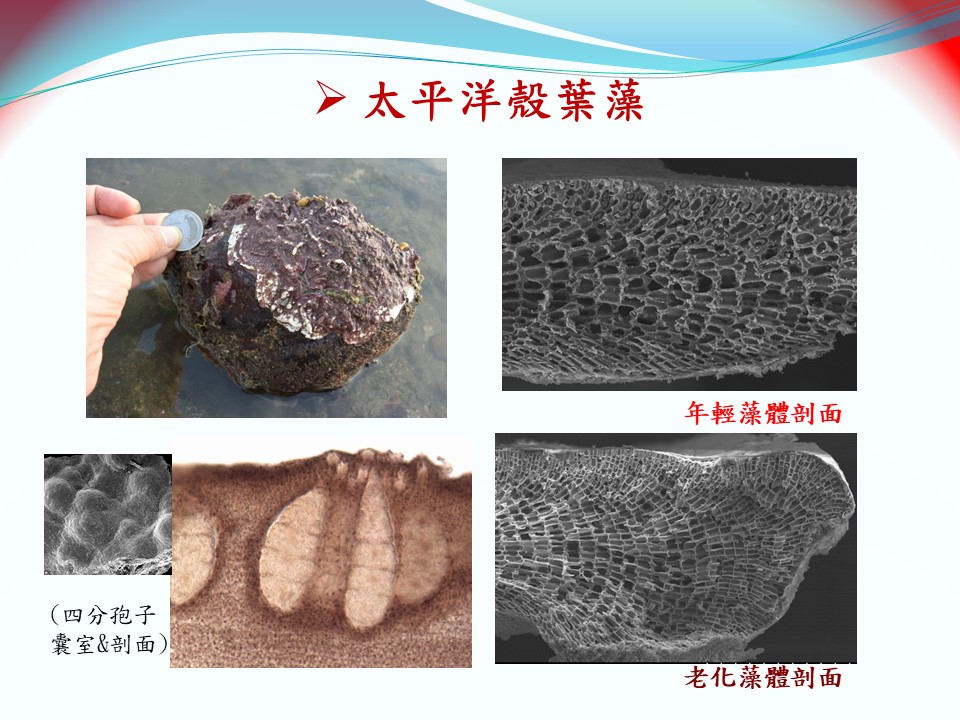
太平洋殼葉藻(上):
拉丁學名:Crustaphytum pacificum L.-C.Liu & S.-M.Lin in Liu et al., 2018
模式種產地:桃園新屋藻礁
棲地:桃園藻礁區低潮位的礁岩或鵝卵石上,亦可在2-3 m深的亞潮帶區域發現。
地理分布:目前僅紀錄於西太平洋的台灣桃園海域(觀新藻礁、大潭藻礁、白玉藻礁)以及法屬新喀里多尼亞島。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
太平洋殼葉藻(下):
形態特徵:
藻體由薄殼狀的鈣化小葉片組成,小葉片可為不規則碎片狀或由具波浪邊緣的橢圓形葉片,葉片長約4~10 cm,寬約3~5 cm。鈣化藻體表面呈現深紫色或深紅色,藻體背面由5-10層或以上的皮層細胞(含有行光合作用色素)組成,藻體腹面(向地面)緊貼著礁岩基質或其他鈣化生物生長。因此在野外觀察殼葉藻的藻體,可看到的鈣化小葉片邊緣呈波浪狀,葉片邊緣不緊貼於礁岩上,藻體表面呈現出碎浪波紋狀。鈣化藻體厚度約100~300 μm,約35~45層細胞厚。鈣化葉片縱切面可觀察到葉片下層的髓質細胞由葉片內部向外呈放射狀生長。四分孢子體世代具有多孔的四分孢子囊生殖窩,具平頂狀的四分孢子囊生殖窩會凸出藻體表面,生殖窩內有多個帶狀分裂的四分孢子囊(目字形)。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
玫瑰哈維石藻(上):
學名:Harveylithon rosea L.-C.Liu & S.-M.Lin in Liu et al., 2018
模式種產地:桃園新屋藻礁。
棲地:較常出現在藻礁區的礁岩或鵝卵石上,亦可在1~2 m 深的亞潮帶藻礁區域發現。
地理分布:廣泛分布於台灣北部沿海各地,包括桃園市、新北市和基隆市。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
玫瑰哈維石藻(下):
形態特徵:
無節的鈣化藻體,外觀近圓形或橢圓形殼狀,長約2~5 cm,寬約3~4 cm,藻體邊緣常呈淺裂狀或略有波浪弧狀。藻體背腹分明。藻體背面為照光面藻體背面由15-30層或以上的皮層細胞(含有行光合作用色素)組成,背面的皮層細胞中含有可行光合作用的色素,使藻體表面呈現粉紅色或玫瑰色。腹面為背光面,腹面藻體緊貼著基質(礁岩或其他鈣化生物)生長。藻體厚度約300~500 μm,約25~50層細胞厚。藻體縱切面可觀察到髓質明顯鈣化葉片縱切面可觀察到葉片下層的髓質細胞由葉片內部向上呈半放射狀生長。四分孢子體世代具有單孔的四分孢子囊生殖窩,生殖窩不凸出或微微凸出於藻體表面。生殖窩內有多個帶狀分裂的四分孢子囊。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021

波緣膨石藻(上):
學名:Phymatolithon margoundulatus L.-C.Liu & S.-M.Lin in Liu et al., 2018
模式種產地:桃園新屋藻礁。
棲地:較常出現在藻礁區的礁岩或鵝卵石上,廣泛分布於高潮位藻礁區,亦可由高潮位至1~3 m深的亞潮帶藻礁區域發現。
地理分布:目前僅紀錄於台灣桃園海域(觀新藻礁、大潭藻礁、白玉藻礁)以及北海岸(石門—新北市)。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
波緣膨石藻(下):
形態特徵:
無節的鈣化藻體,外觀近扇形平面藻體或扁平不連續狀的玫瑰花座形,長約2~4 cm,寬約2~3 cm,藻體邊緣常呈淺裂狀或略有波浪弧狀。藻體背腹分明。藻體背面為照光面,藻體背面由10-15層或以上的皮層細胞(含有行光合作用色素)組成,背面的皮層細胞中含有可行光合作用的色素,使藻體表面呈現紫紅色到深紅色,藻體邊緣多為白色或淺粉色。腹面為背光面,腹面藻體緊貼著基質(礁岩或其他鈣化生物)生長。藻體厚度約100~250 μm,約35~45層細胞厚。藻體縱切面可觀察到髓質細胞由葉片內部向上呈半放射狀生長。四分孢子體世代具有多孔的四分孢子囊生殖窩,生殖窩以半球狀凸出於藻體表面。生殖窩內有多個帶狀分裂的四分孢子囊。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021

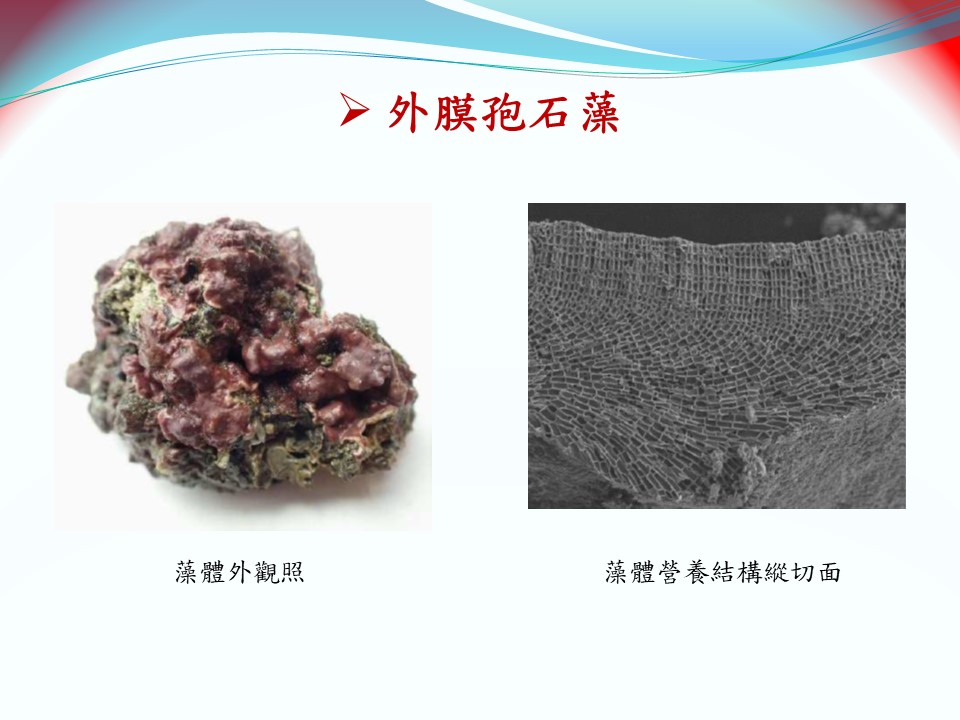
外膜孢石藻(上):
學名:Sporolithon episporum (M.Howe) E.Y.Dawson, 1960
模式種產地:巴拿馬
棲地:廣泛分布於藻礁區中低潮位的潮間帶礁岩上。
地理分布:廣泛分布於印度洋(南非、印尼、西澳)、西太平洋(臺灣、日本、紐西蘭巴布亞新幾內亞、南太平洋群島)、大西洋(巴西)以及加勒比海海域(哥斯大黎加與巴拿馬)。台灣主要分布於桃園海域(觀新藻礁、大潭藻礁、白玉藻礁)、澎湖群島海域(吉貝、尖山)以及屏東墾丁國家公園(眺石)。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
外膜孢石藻(下):
形態特徵:
無節的鈣化藻體,扁平至凸瘤狀的藻體緊貼於礁岩上生長,藻體直徑可達5 cm以上。藻體表面平滑。藻體背腹分明,背面為照光面,藻體背面皮層細胞可達40層以上,背面的皮層細胞中含有可行光合作用的色素,使藻體表面呈現紫紅色至紅棕色,藻體有白色大波浪狀邊緣。腹面為背光面,腹面藻體緊貼著基質(礁岩或其他鈣化生物)生長。藻體邊緣外觀與太平洋殼葉藻相似,然而外膜孢石藻邊緣緊貼,太平洋殼葉藻邊緣常不緊貼於基質上。藻體厚度可達800 μm以上(約85~120層細胞)。藻體縱切面可觀察到髓質明顯有鈣化葉片縱切面可觀察到葉片下層的髓質細胞由葉片內部向上呈半放射狀生長。成熟四分孢子體具有許多單孔的鈣化窩,而且鈣化窩不凸出於藻體表面。每個鈣化窩只產生一個十字型分裂四分孢子囊。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021
瑪格麗特石葉藻:
學名:Lithophyllum margaritae (Hariot) Heydrich, 1901
模式種產地:墨西哥
棲地:廣泛分布於藻礁區的礁岩或鵝卵石上,亦可在1~2 m 深的亞潮帶藻礁區域發現。
地理分布:廣泛分布於太平洋(加利福尼亞灣、台灣)、西大西洋(巴西、墨西哥)。台灣主要分布於桃園海域(觀新藻礁、大潭藻礁、白玉藻礁);東北角海域的龍洞灣(新北市)。
形態特徵:
無節的鈣化藻體,外觀為扁平狀殼狀,藻體中心少有瘤狀突起。表面呈現淡粉色、亮紅色或紅棕色。長約5~6 cm,寬約6~8 cm,藻體邊緣為全緣。藻體厚度約300~1000 μm,藻體可達160層以上細胞厚。藻體縱切面可觀察到藻體由基部貼緊基質生長的基底絲與垂直生長的直立絲組成。四分孢子體世代具有單孔的四分孢子囊生殖窩,生殖窩微凸出於藻體表面。生殖窩內有多個帶狀分裂的四分孢子囊。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021

薩摩亞哈維石藻(上):
學名:Harveylithon samoënse (Foslie) A.Rösler, Perfectti, V.Peña & J.C.Braga,2016
模式種產地:薩摩亞
棲地:潮間帶低潮位的礁岩或鵝卵石上,亦可在2-3 m 深的亞潮帶區域發現。
地理分布:廣泛分布於印度洋及西太平洋,台灣目前記錄於桃園海域(觀新藻礁、大潭藻礁、白玉藻礁)。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,20
薩摩亞哈維石藻(下):
形態特徵:
無節的鈣化藻體,外觀為不規則橢圓形殼狀,長約2~5 cm,寬約3~4 cm,藻體邊緣常呈淺裂狀或略有圓弧狀突出。藻體背腹分明,厚度可達280μm 以上。藻體背面為照光面藻體背面由60-75 層或以上的皮層細胞(含有行光合作用色素)組成,且具扁平狀表皮細胞。背面的皮層細胞中含有可行光合作用的色素,使藻體表面呈現粉橘色或紅橘色。腹面為背光面,腹面藻體緊貼著基質(礁岩或其他鈣化生物)生長。四分孢子體世代具有單孔的四分孢子囊生殖窩,而且生殖窩內有多個帶狀分裂的四分孢子囊。
資料來源:
「110年-111年藻礁生態系調查計畫(1/2)成果報告書」-海保署,2021

生態影音

2022/04/09
大潭海岸貝殼沙

2022/04/09
白玉海岸貝殼沙

2022/12/12
大潭G2生態調查魚群